 Profile
Profile Settings
Settings Refer your friends
Refer your friends Sign out
Sign out





The tapetum is a specialised layer of nutritive cells placed between the sporogenous tissue and the anther wall of flowering plants. Tapetum is critical for pollen grain nutrition and development, as well as a source of precursors for the pollen coat. Cells are often larger and contain several nuclei. As sporogenous cells divide, the nuclei of tapetal cells divide as well. At times, this mitosis is abnormal, resulting in the multinucleation of a large number of mature tapetum cells. Occasionally, polyploidy and polytenity are observed. The tapetum’s extremely large nuclear composition aids in the provision of nutrients and regulatory chemicals to developing pollen grains. This is accomplished using the following processes:
Endomitosis
Normal mitosis without cytokinesis
Restitution nuclei formation
Endoreduplication
Tapetum aids in the construction of the pollenwall, the delivery of nutrients to the inner side of the anther, and the manufacture of the callase enzyme required for the separation of microspore tetrads.
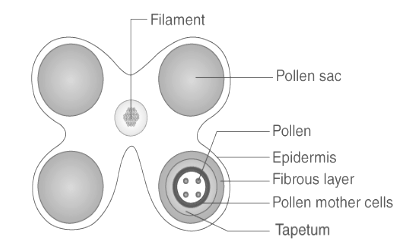
Structure of Tapetum
The tapetum is a dense single-celled nutritive layer that lies under the epidermis and endodermis. Their cells begin as diploid but can become polyploid as a result of aberrant cell division. They are primarily responsible for absorbing nutrients from the middle layer and supplying them to the microsporocytes. Additionally, tapetum cells are capable of secreting enzymes and hormones. In the adult anther, the tapetum layer is almost completely absent.
Types of Tapetum
The tapetum is classified into two distinct categories based on its behaviour:
1. Amoeboid 2. Secretory Tapetum.
1. Amoeboid or Invasive or Periplasmodial Tapetum
Alisma, Butomus, Tradescantia, and Typha all have this type of tapetum, in which the tapetal cells combine to create a tapetal periplasmodium. The united tapetal cells’ protoplasts migrate to the locule, where they surround the pollen mother cells or forming pollen grains. This transfer of the protoplast into the locule may occur during meiotic prophase or may be postponed until the tetrad stage.
Tradescantia tapetal cells become increasingly vacuolated during the pre-meiotic stage of sporogenous cell growth, and vesicles, probably formed from dictyosomes, begin to exit the cell. These vesicles contain enzymes that accelerate and complete the degradation of the tapetal cell wall. Simultaneously, the pollen mother cells undergo meiotic divisions to produce microspores.
A wave of lysis propagates across the anther locules, dehydrating the tapetal cells. The overall result of these activities is an anther cavity containing microspore tetrads that has been invaded by tapetal protoplast that now contains a substantially higher concentration of polysaccharide material.
The new peri plasmodial tapetal cytoplasm undergoes additional remodelling at the end of pollen mother cell meiosis. The tapetal protoplasts’ callases gradually detach the microspore tetrads of their callose covering, and the long amoeboid-like tapetal cells penetrate between the freshly freed immature pollen grains.
The tapetal cell metabolism then switches from carbohydrate production and degradation to lipid synthesis. Within the plastids, lipid globules form and are gradually extruded into the tapetal cytoplasm.
Around the time of final mitosis, the tapetal cytoplasm becomes increasingly vacuolated, indicating the onset of a second phase of polysaccharide production. Finally, alterations in the anther cuticle and connective tissue result in significant breakdown of the tapetal cytoplasm, which is deposited on the surface of the pollen grains as a thin tryphine (complex mixture of hydrophilic chemicals) layer during anthesis.
Mepham and Lane (1969) investigated the ultrastructure of this Tradescantia tapetum variety and demonstrated that it contained an organised and functional structure. The peri plasmodium infiltrates between the microspore mother cells and finally engulfs each pollen mother cell, where it plays a critical function in microspore development.
The periplasm sodium cell organelles undergo rearrangement and nuclear division during development. At the premeiotic stage of sporogenous tissue, hydrolytic enzymes released by the dictyosomes in the tapetal cells degrade the tapetal cell walls.
Hydrolytic enzymes may also penetrate and digest the walls of some sporogenous cells located within the anther locule. Following completion of meiosis, the callose wall around the spore degenerates enzymatically from the tapetal Plasmodium, and as a result, the spores are bathed in tapetal cytoplasm before to anthesis.
2. Glandular or Secretory Tapetum
It is the most prevalent type of tapetum in angiosperms, in which the cells retain their original location and gradually degrade. Pollenkitt and tryphine are two of the substances secreted by the degenerating tapetum.
Echlin and Godwin (1968) detailed the events that occurred in the secretory tapetal cells of Helleborus foetidus from the premeiotic stage until pollen maturation, which are summarised below:
The cell wall is relatively thin, consisting of a primary wall with a low cellulose content and a middle lamella. The cytoplasm is composed of mitochondria, plastids, pro-Ubisch bodies, and dictyosomes.
Prior to meiosis, the tapetal cell wall thickens and the cytoplasm gets thicker as the number of ribosomes and pro-Ubisch bodies increases. However, the wall confronting the locule is thickened irregularly.
During meiosis, the number of pro-Ubisch bodies increases in tandem with the nuclear size.
The pro-Ubisch bodies are surrounded by a radiative zone of ribosomes during the tetrad stage. After being detached from the tetrad, the pro-Ubisch bodies penetrate through the tapetal cell membrane and are extruded into the space between the membrane and the cell wall, where they are instantly coated with sporopollenin and are dubbed Ubisch bodies.
The tapetal cell’s inner tangential wall appears thinner, and a space between the cell membrane and the wall appears. This is followed by the wall disintegrating and a new membrane around the tapetal protoplast forming along the thecal phase.
The tapetal membrane is the name given to this layer. This layer is resistant to acetolysis, is studded with Ubisch granules, and forms a sac around the developing pollen grains in the anther locule.
Function of Tapetum
Tapetum supplies nutrients for the pollen grains as they mature.
Additionally, they serve as a source of precursors for the pollen wall or pollen coat.
It carries nutrients to the anthers.
Additionally, pollenkitt is generated by the tapetal cells that surround the microsporocytes. This pollenkitt is in charge of feeding and pollen production.
Tapetal cells serve as precursors to sporopollenin. Pollen grains’ exterior walls include a substance called sporopollenin, which protects the pollen from harsh environmental influences.
Tapetum excretes Ubisch bodies, which contribute to the pollen grain membrane’s thickness. These ubisch entities, also known as orbicules, are a component of sporopollenin.
Additionally, it produces the callase enzyme, which degrades the callose molecules required to connect four pollen (pollen tetrad). Thus, they enable the separation of pollen from a tetrad.
Conclusion
There are two distinct forms of tapetum: secretory (glandular) and plasmodial (amoeboid). A layer of tapetal cells surrounds the anther locule in the secretory type, whereas in the plasmodial type, the tapetal cell walls disintegrate and their protoplasts combine to create a multinucleate plasmodium. In Canna, a third, less common variety of tapetum has been described: the invasive non-syncytial tapetum, in which the tapetal cell walls degrade to penetrate the anther locule but do not fuse to create a plasmodium.
Acorales, the first branching group of monocots, contains a secretory tapetum, whereas Alismatales, the other alismatidae clade, is largely plasmodial. Among the late branching clades, the lilioid monocots are almost exclusively secretory, but the commelinid monocots exhibit a range of tapetal patterns.
